
Table of Contents
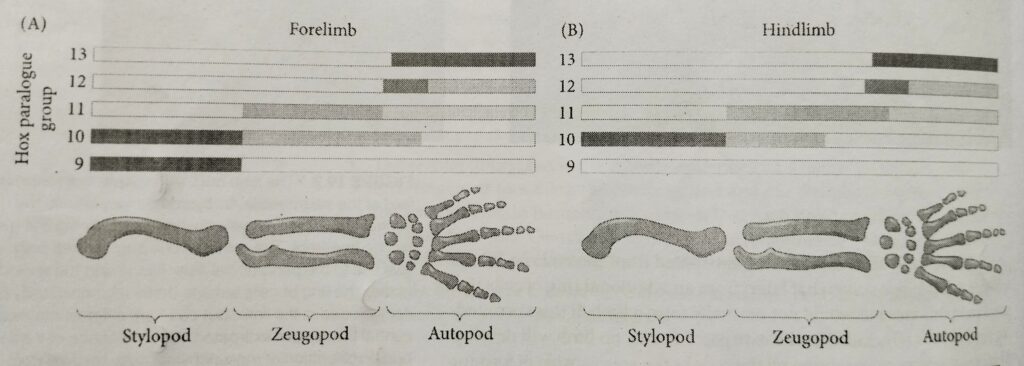
The bones of any tetrapod limbs (arm or leg, wing or flipper) consist of a Proximal stylopod (humerus/ femur) adjacent to the body wall, a zeugopod (radius-ulna/tibia-fibula) in the middle region and a distal autopod (carpals-fingers/ tarsals -toes).
The Tetrapod limbs bud
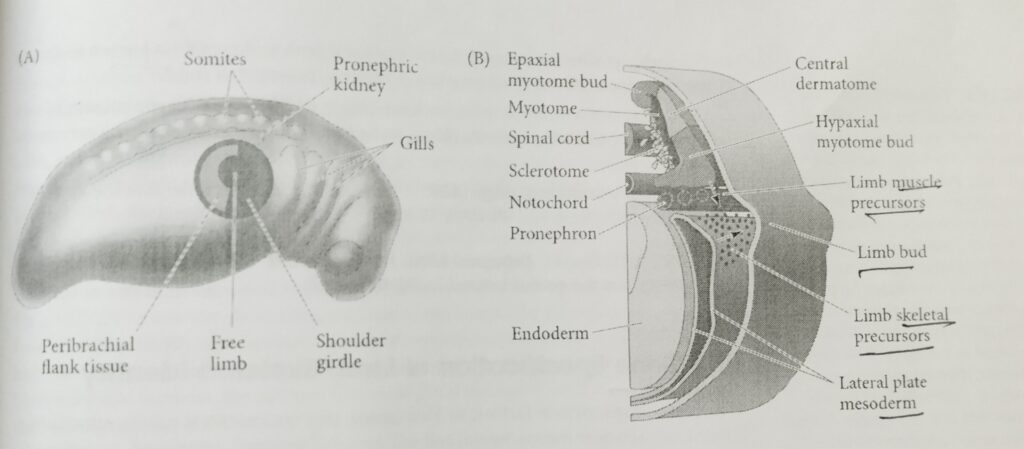
The first visible sign of limb development is the formation of bilateral bulges called limb buds at the presumptive forelimb and hindlimb locations.
Adjacent to it are the cells that will form the peri brachial (around the limb)flank tissue and the shoulder girdle. The larger region, representing all the cells in the area capable of forming a limb on their own, is the limb field.
lateral plate mesenchyme cells migrate within the limb fields to form the limb muscle precursor cells. This accumulating heterogenous population of mesenchymal cells proliferates under the ectodermal tissue, generating the limb bud.
Hox gene specification of tetrapod limb
The 5′ (AbdB-like) portions (paralogues 9-13) of the Hoxa and Hoxd gene complexes appear to be active in the limb buds of mice.
Here, Hox9 and Hox10 paralogues specify the stylopod, Hox 11 paralogues specify the zeugopod, Hox12, and Hox13 paralogues specify the autopod.
Induction of Tetrapod limbs bud
This process is divided into four stages:
- making mesoderm permissive for limb formation
- specifying forelimb and hindlimb
- inducing epithelial-to-mesenchymal transition
- establishing two positive feedback loops for limb bud formation.
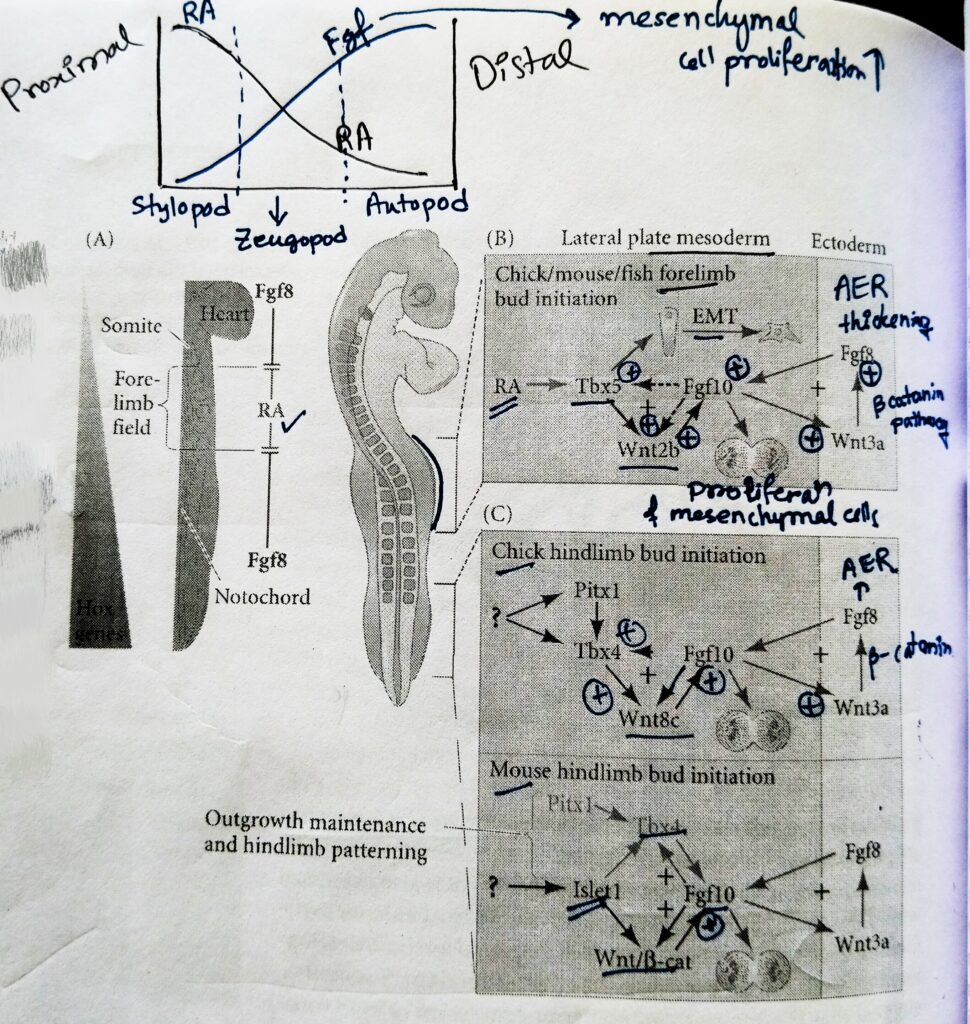
1) Making mesoderm permissive for tetrapod limbs formation:
- The relationship between retinoic acid (RA) and Fgf8 during somitogenesis is antagonist.
- The Fgf8 is expressed by the caudal progenitor zone located just posterior to the forelimb field, whereas RA is generated more anteriorly in somites and anterior presomitic mesoderm; relevant for forelimb development, however, Fgf8 is also expressed within the heart lateral plate mesoderm located just anterior to the forelimb field.
- In contrast, RA is present throughout the somitic regions of the trunk adjacent to the forelimb field, where it is required to repress trunk Fgf8 expression and thus promote forelimb bud initiation.
- Targeted loss of RA synthesis in the vertebrate forelimb results in an expansion of Fgf8 expression towards the presumptive forelimb field. Which leads to the reduction in the Tbx5 expression, and the failure to form forelimb bud.
- In the absence of Fgf8, the lateral plate mesoderm is permissive for forelimb bud formation and development.
- Although RA-Fgf8 antagonism plays a role during forelimb initiation and the early stages of somitogenesis. RA inhibits hindlimb development. Loss of RA synthesis in mice does not effect hindlimb bud formation.
2) Forelimb and hindlimb specification by Tbx5 and Islet1, respectively:
- Early specification of forelimb and hindlimb identity begins in the limb fields prior to tetrapod limbs bud formation through the expression of particular transcription factors.
- In mice, the genetic encoding Tbx5 is transcribed in the limb fields of the forelimbs, while the genes encoding Islet1, Tbx4, and Pitx1 are expressed in presumptive hindlimbs.
- Downstream of the regulatory function of these transcription factors of Fgf10, the primary inducer of limb bud formation through the initiation of cell shape changes and proliferation of outgrowth.
- When Fgf10- secreting beads were used to induce an ectopic limb between the chicken hindlimb and forelimb buds, this type of limb produced was determined by which Tbx protein was expressed.
- Limb buds induced by placing FGF beads close to the hindlimb (opposite somite 25) expressed Tbx4 and become hindlimb.
- Limb buds induced close to the forelimb (opposite somite 17) expressed Tbx5 and developed as forelimb (wings).
- Limb buds induced in the centre of the flank tissue expressed Tbx5 in the anterior portion of the limb and Tbx4 in the posterior portion, these limbs developed as chimeric structures, with the anterior resembling a forelimb and the posterior resembling a hindlimb.
- More recent investigations have revealed two additional transcription factors involved in initiating the hindlimb: Pitx1 and Islet1. Indeed, misexpressions of Pitx1 in the mouse forelimb causes its muscles, bones, and tendons to develop into ones that look like those of a hindlimb.
- Additionally, Pitx1 protein activates hindlimb-specific genes in the forelimb, including Hoxc10 and Tbx4. Interestingly, a mutation in the human PITX1 gene that causes a haploinsufficiency in Pitx1 protein leads to the bilateral “club foot” phenotype.
- Islet1 is a homeodomain transcription factor, which is transiently expressed in the hindlimb field before Fgf10 expression and leg bud formation in mice.
- Transcriptional regulation of Islet1 and Pitx1 are independent of each other’s function, as are their roles in hindlimb development.
3) Induction of epithelial-to-mesenchymal transition by Tbx5:
- Research by Clifford Tabin’s lab has demonstrated that the epithelial cells making up the mesoderm of the early somatopleure undergo an epithelial-to- mesenchymal transition (EMT) specially in the limb fields before any signs of such cell behaviour is observed in the mesoderm of flank regions.
- At least in the case of the forelimb, Tbx5 knockout mice show a significant loss of limb bud mesenchyme, suggesting that Tbx5 is a major regulator of EMT in the forelimb field.
4) Establishing two positive feedback loops for limb bud formation by Fgf-Wnt:
- Through the upregulation of Tbx5 in the forelimb and Islet1 in the hindlimb, mesenchymal cells commit towards limb bud development and secrete the paracrine factor Fgf10.
- Fgf10 provides the signal to initiate the propagate the limb- forming interactions between ectoderm and mesoderm, these signaling interactions directly promote limb bud formation and growth.
- Limb development is a prolonged process of outgrowth, and Fgf10 possesses the morphogenetic power to induce limb formation. A bead containing Fgf10 placed ectopically beneath the flank ectoderm can induce extra limbs to form.
- Among the downstream targets of Fgf10 signalling are Wnt/β- catenin and the transcription factors it initiates, which perpetuate Fgf10 signalling.
- In the presumptive forelimb field, for example, Tbx5 induces Wnt2b, which then upregulates Fgf10, which in turn positively feeds back to maintain activation of both Wnt2b and Tbx5.
- Fgf10 is secreted by the tetrapod limbs field mesenchyme induces the overlying ectoderm to form apical ectodermal ridge (AER). The AER runs along the distal margin of the limb bud and will become a major signaling center for developing limbs.
- Fgf10 is capable of inducing the AER in the competent ectoderm between the dorsal and the ventral side of embryo.
- Fgf10 stimulates Wnt3a (Wnt3a in chicks; Wnt3 in humans and mice) in the prospective limb bud surface ectoderm. The Wnt protein acts through the canonical β-catenin pathway to induce Fgf8 expression in the ectoderm. This relay of signaling represents a foundational stage in the formation of limb buds, because once Fgf8 is made in the surface ectoderm, the surface ectoderm elongates to physically become the AER.
- One of the main functions of AER is to tell the mesenchyme cells directly beneath it to continue making Fgf10.
- In this way, a second positive feedback loop is created wherein mesodermal Fgf10 tells the surface ectoderm to continue to make Fgf8, and the surface ectoderm continues to tell the underlying mesoderm to make Fgf10.
Diagram of Tetrapod limbs field initiation:
The note of Tetrapod limbs development is made from the development biology book by Gilbert.
Other related notes:
- Patterns of cleavage: https://thebiologyislove.com/patterns-of-cleavage/
- Gamete fusion and prevention of polyspermy: https://thebiologyislove.com/gamete-fusion-the-prevention-of-polyspermy/
Facebook link: https://www.facebook.com/share/p/oV4NN56FM4uLE8XJ/?mibextid=oFDknk